根(4)
前面已经讨论了根尖的结构和它的发育情况,由根尖的顶端分生组织,经过分裂、生长、分化而形成成熟的根,这种植物体的生长,直接来自顶端分生组织的衍生细胞的增生和成熟,整个生长过程,称为初生生长(primary growth)。初生生长过程中产生的各种成熟组织属于初生组织(primary tissue),它们共同组成根的结构,也就是根的初生结构(primary structure)。因此,在根尖的成熟区作一横切面,就能看到根的全部初生结构,由外至内为表皮、皮层和维管柱三个部分(图3-9)。
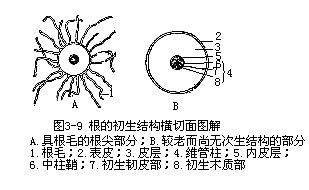
(一)表皮
根的成熟区的最外面具有表皮,是由原表皮发育而成,一般由一层表皮细胞组成,表皮细胞近似长方柱形,延长的面和根的纵轴平行,排列整齐紧密,和植物体其他部分一般的表皮组织相似。但根的表皮细胞壁薄,角质层薄,不具气孔,部分表皮细胞的外壁向外突起,延伸成根毛。成熟的根毛直径5—17μm,长 80—1500μm,因种而异。少数植物,如洋葱无根毛。根的这些特征是和它的吸收、固着等作用密切有关。根的表皮,一般是由一层活细胞组成的,但也有例外,在热带的兰科植物和一些附生的天南星科植物的气生根中,表皮是多层的,形成所谓根被(velamen)。根被是由紧密排列的死细胞组成的鞘,这些死细胞的壁由带状或网状增厚来加固,壁上有许多初生纹孔场。当空气干燥时,这些细胞充满着空气;当降雨时,它们就充满了水。当根由水气饱和时,根被也有气体交换的功能。有的科学家在兰科植物的研究上,认为根被的主要作用是机械的保护作用和防止皮层中过多水分的丧失,而过去认为根被具吸收水分的作用,尚待证实。
(二)皮层
皮层是由基本分生组织发育而成,它在表皮的内方占着相当大的部分,由多层薄壁细胞组成,细胞排列疏松,有着显著的胞间隙。皮层最外的一层细胞,即紧接表皮的一层细胞,往往排列紧密,无间隙,成为连续的一层,称为外皮层(exodermis)。当根毛枯死,表皮破坏后,外皮层的细胞壁增厚并栓化,能代替表皮起保护作用。有些植物的根如鸢尾,外皮层为多层细胞组成。
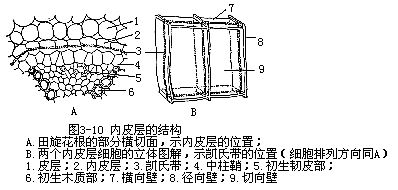
皮层最内的一层,常由一层细胞组成,排列整齐紧密,无胞间隙,称为内皮层(endoder-mis)。内皮层细胞的部分初生壁上,常有栓质化和木质化增厚成带状的壁结构,环绕在细胞的径向壁和横向壁上,成一整圈,称凯氏带(Casparian strip,图3-10),凯氏带在根内是一个对水分和溶质有着障碍或限制作用的结构。凯氏带形成后,内皮层的质膜与凯氏带(即被木质和栓质沉积的细胞壁部分)之间有极强的联系,水分和离子必须经过这个质膜,才能进入维管柱,这里也就有着选择。在电子显微镜下观察到质壁分离的细胞中,质膜紧贴着凯氏带区,只有这个区以外的质膜才分离开(图3-11)。另外,现代科学技术如放射自显影等的研究已证明,由于凯氏带的存在,皮层胞壁间的运输只到凯氏带处,不能超越,而根尖较幼部分的内皮层,由于尚未充分分化和凯氏带尚未形成,细胞壁间的运输仍可直接和木质部相通,都能说明内皮层是有限制作用的结构。在单子叶植物根中,内皮层的进一步发展,不仅径向壁及横向壁因沉积木质和栓质而显著增厚,而且在内切向壁(向维管柱的一面)上,也同样地因木质化和栓质化而增厚,只有外切向壁仍保持薄壁(图3-12)。增厚的内切向壁上有孔存在,以便使通过质膜的中的细胞质某些溶质,能穿越增厚的内皮层。另外,少数位于木质部束处的内皮层细胞,仍保持初期发育阶段的结构,即细胞具凯氏带,但壁不增厚的,称为通道细胞(passage cell),起着皮层与维管柱间物质交流的作用。


(三)维管柱
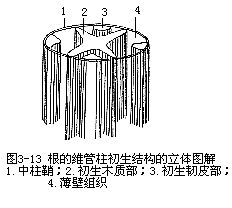
维管柱是内皮层以内的部分,结构比较复杂,包括中柱鞘(pericycle)和初生维管组织,有些植物的根还具有髓(pith),由薄壁组织或厚壁组织组成。
中柱鞘是维管柱的外层组织,向外紧贴着内皮层。它是由原形成层的细胞发育而成,保持着潜在的分生能力,通常由一层薄壁细胞组成,也有由两层或多层细胞组成的,有时也可能含有厚壁细胞。维管形成层(部分的)、木栓形成层、不定芽、侧根和不定根,都可能由中柱鞘的细胞产生。
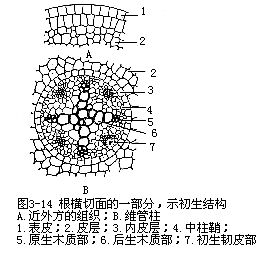
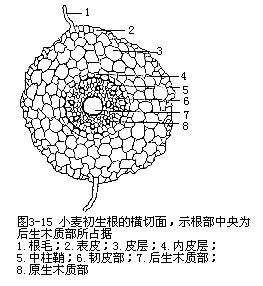
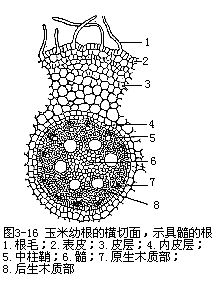
根的维管柱中的初生维管组织,包括初生木质部(primary xylem)和初生韧皮部(pri-mary phloem),不并列成束,而是相间排列,各自成束(图3-13)。由于根的初生木质部在分化过程中,是由外方开始向内方逐渐发育成熟,这种方式称为外始式(exarch),这是根发育上的一个特点。因此,初生木质部的外方,也就是近中柱鞘的部位,是最初成熟的部分,称为原生木质部(protoxylem),它是由管腔较小的环纹导管或螺纹导管组成。渐近中部,成熟较迟的部分,称为后生木质部(metaxylem),它是由管腔较大的梯纹、网纹或孔纹等导管所组成。由于初生木质部的发育是外始式,因此,外方的导管最先形成,这就缩短了皮层和初生木质部间的距离,从而加速了由根毛所吸收的物质向地上部分运输(图3-14)。在根的横切面上,初生木质部整个轮廓呈辐射状,而原生木质部构成辐射状的棱角,即木质部脊(xylem ridge)。不同植物的根中,木质部脊数是相对稳定的,例如烟草、油菜、萝卜、胡萝卜、芥菜、甜菜等是2束,紫云英、豌豆等是3束,蚕豆、落花生、棉、向日葵、毛茛、蓖麻等是4束,有时5束,茶、马铃薯是5束,葱是6束,多于6束的,如葡萄、菖蒲、高粱、棕榈、鸢尾、玉米、水稻、小麦等。植物解剖学上,依根内木质部脊数的不同,把根分别划分成二原型(diarch)、三原型(triarch)、四原型(tetrarch)、五原型(pentarch)、六原型(hexarch)和多原型(polyarch)等。初生木质部束也常发生变化,同种植物的不同品种中,例如茶有5束、6束、8束、甚至12束等。同一株植物的不同根上,可能出现不同束数,如落花生主根为4束,而侧根有时出现2束。近年来的研究发现,在离体培养的根中,培养基中生长素吲哚乙酸(IAA)的含量高低,可以影响木质部脊数。初生木质部的结构比较简单,主要是导管、管胞,也有木纤维和木薄壁组织。初生韧皮部发育成熟的方式,也是外始式,即原生韧皮部(protophloem)在外方,后生韧皮部(meta-phloem)在内方。初生韧皮部束数在同一根内,与初生木质部脊数相等,它与初生木质部脊相间排列,即位于初生木质部二束之间。初生韧皮部由筛管和伴胞组成,也含有韧皮薄壁组织,有时还有韧皮纤维,如锦葵科、豆科、番荔枝科植物。初生木质部和初生韧皮部之间,也分布着薄壁组织。一般植物根的中央部分往往由初生木质部中的后生木质部占据(图3-15),如果中央部分不分化成木质部,就由薄壁组织或厚壁组织形成髓。多数单子叶植物,以及双子叶植物中的有些草本植物和多数木本植物的根,存在着髓。蚕豆、落花生、玉米、高粱等具髓的根,它们的初生木质部和初生韧皮部,在髓的外围也作相间排列的方式(图3-16)。
免责声明:本文来源于网络,文中有些文字或数据已经过期失效,仅供学习备课参考!
电脑版地址:http://www.cgzdl.com/shuku/244/9744.html
手机版地址:http://m.cgzdl.com/shuku/244/9744.html





-lp.jpg "甘肃省兰州市旅游地图高清版")

