绿藻门(Chlorophyta)
第八节 绿藻门(Chlorophyta)
一、绿藻门的一般特征
(一)形态与构造
绿藻门植物体的形态是多种多样的,有单细胞、群体、丝状体和叶状体。少数单细胞和群体类型的营养细胞前端有鞭毛,终生能运动。绝大多数绿藻的营养体不能运动,只在繁殖时形成的游动孢子和配子有鞭毛,能运动。
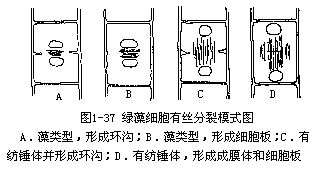
绿藻细胞有细胞壁,分两层,是原生质体分泌的。内层主要成分为纤维素,外层是果胶质,常常粘液化。在原始类型中,细胞里充满原生质,或在原生质中只形成很小的液泡。在高级类型中,像高等植物一样,中央有1个充满着细胞液的大液泡。气生类型的藻类细胞中,则无中央大液泡。绿藻细胞中的载色体和高等植物的叶绿体结构类似,电子显微镜下观察,有双层膜包围,光合片层为3—6条叠成束排列。载色体中还含有DNA,但不是所有载色体都有,据分析,伞藻属(Acetabularia)细胞中的载色体,仅20—25%含DNA。载色体所含的色素也和高等植物相同,主要色素有叶绿素a和b、α-胡萝卜素和β-胡萝卜素以及一些叶黄素类。在载色体内通常有1至数枚蛋白核。同化产物是淀粉,其组成与高等植物的淀粉类似,也是由直链淀粉和支链淀粉组成。淀粉多贮存于蛋白核周围。有时也贮存蛋白质和油。细胞核1至多数,通常位于靠壁的原生质中。单核种类细胞核常位于中央,悬在原生质丝上(如水绵属)。细胞是有丝分裂,有两个基本类型:1种类型是在细胞分裂中期核膜不消失,或仅在极端形成孔,末期纺锤体消失,仅有与纺锤体纵轴垂直排列的纺锤丝,末期两子核相距很近,称此种有丝分裂为藻类型,是绿藻纲植物分裂方式,两子核相距较近,胞质以环沟和细胞板的方式进行分裂;另1种类型是细胞有丝分裂的中期核膜消失,有纺锤体,末期形成成膜体(phragmoplast),两子核相距较远。细胞有丝分裂出现成膜体是轮藻纲和陆生绿色植物的特征(图1-37)。运动细胞具两条或四条顶生等长鞭毛。电子显微镜下观察,鞭毛是由9+2条轴丝组成。轴丝几乎全被鞭毛鞘包围,仅末梢裸露。鞭毛鞘上没有羽状鞭茸结构,故称此种鞭毛为尾鞭型。
(二)繁殖
绿藻的繁殖有营养繁殖、无性生殖和有性生殖。
1.营养繁殖 群体、丝状体以细胞分裂来增加细胞的数目。大的群体和丝状体常由于动物摄食、流水冲击等机械作用,使其断裂;或由于丝状体中某些细胞形成孢子或配子,在放出配子或孢子后的空细胞处断裂;或由于丝状体中细胞间胶质膨胀分离,而形成单个细胞或几个细胞的短丝状体[如裂丝藻属(Stichococcus)],断裂的每一小段都可发育成新的藻体。某些单细胞绿藻遇到不良环境时,细胞多次分裂形成胶群体,环境好转时,每个细胞又可发育成1个新的植物体。
2.无性生殖 无性生殖可形成游动孢子(zoospore)或静孢子(aplanospore)。游动孢子无壁,其他构造和衣藻属的细胞相似。形成游动孢子的细胞和普通营养细胞没有区别。有些种的藻体,全体细胞都可产生游动孢子,但是,群体类型的藻体,不是所有细胞都同时形成游动孢子。有些藻类仅限于一定的细胞中产生游动孢子。形成游动孢子时,细胞内原生质体收缩,形成1个游动孢子,或经过分裂形成多个游动孢子,其数目是2的次方。游动孢子多在夜间形成,黎明时放出,或在环境突变时形成游动孢子。游动孢子放出后,游动一个时期,缩回或脱掉鞭毛,分泌1层壁,成为1个营养细胞,继而发育为新的植物体。有些藻类以静孢子进行生殖。静孢子无鞭毛,不能运动,有细胞壁。另有1种静孢子,在形态上与母细胞相同,称似亲孢子(autospore)。在环境条件不良时,细胞原生质体分泌厚壁,围绕在原生质体的周围,并与原有的细胞壁愈合,同时细胞内积累大量的营养物质,形成厚壁孢子,环境适宜时,发育成新的个体。
3.有性生殖 有性生殖的生殖细胞叫配子(gamete)。两个生殖细胞结合形成合子(zygote),合子直接萌发成新个体,或经过减数分裂(reduction pision)形成孢子,并发育成新个体。在形状、结构、大小和运动能力等方面完全相同的两个配子结合,称为同配生殖(isogamy)。在形状和结构上相同,但大小和运动能力不同,大而运动能力迟缓的为雌配子(female gamete),小而运动能力强的为雄配子(male gamete),此两种配子的结合称为异配生殖(anisogamy)。在形状、大小和结构上都不相同的配子,大而无鞭毛不能运动的为卵(egg),小而有鞭毛能运动的为精子(sperm),精卵结合称为卵式生殖(oogamy)。两个没有鞭毛能变形的配子结合,称为接合生殖(conjugation)。
(三)分布
绿藻分布在淡水和海水中,海产种类约占10%,淡水产种类约占90%。有些种类专门生活在谈水中,如鞘藻目(Oedogoniales)和双星藻目(Zygnematales);石莼目(Ulvales)和管藻目(Siphonales)是海产种占优势;丝藻目(Ulotrichales)是淡水种占优势;另外也有不少种生活在半咸水中。海产种多分布在海洋沿岸,往往附着在10米以上浅水中的岩石上。许多海产种有一定的地理分布,这是由于水的温度决定的。淡水种的分布很广,江河、湖泊、沟渠、积水坑中,潮湿的土壤表面,墙壁上,岩石上,树干上,花盆四周,甚至在冰雪上都可找到。它们中部分是沉在水中生活,许多单细胞和群体种类是漂浮在水中,但在海水中没有浮游的绿藻,有的绿藻也可以寄生在动物体内,或者与真菌共生形成地衣。一般淡水种不受水温的限制,大部分分布在世界各地。
绿藻是藻类植物中最大的1门,约有350个属,5000—8000种。分成绿藻纲(Chloro-phyceae)和轮藻纲(Charophyceae)两个纲。我国一般将绿藻纲分为 13个目,即团藻目(Volvocales)、四孢藻目(Tetrasporales)、色球藻目(Chlorococcales)、丝藻目、胶毛藻目(Chaetophorales)、石莼目、溪菜目(Prasiolales)、鞘藻目、刚毛藻目(Cladophorales)、管藻目、管枝藻目(Siphonocladales)、绒枝藻目(Dasycladales)和双星藻目。轮藻纲中只有轮藻目(Charales)。
二、绿藻门的代表植物
(一)衣藻属(Chlamydomonas)
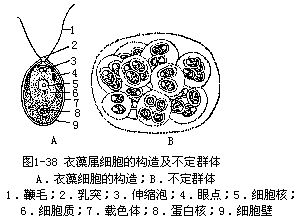
衣藻属是团藻目内单细胞类型中的常见植物。植物体是单细胞,卵形、椭圆形或圆形。体前端有两条顶生鞭毛,有些种在鞭毛着生处有乳头状突起,有的则无,鞭毛是衣藻在水中的运动器官。细胞壁分两层,内层是纤维素的,外层是由果胶质包着。多数种的载色体,其形状如厚底杯形,在基部有1个明显的蛋白核。载色体也有片状的、H形的或星芒状的。细胞中央有1个细胞核。鞭毛基部有两个伸缩泡,伸缩泡是突然收缩的,一般认为是排泄器官。眼点橙红色,位于体前端载色体膜与光合片层之间,由1层或数层油滴构成,油滴中含有类胡萝卜素(图1-38,A)。
无性生殖 衣藻经常在夜间进行无性生殖。生殖时藻体通常静止,鞭毛收缩或脱落变成游动孢子囊。细胞核先分裂,形成4个子核,有些种分裂3—4次,形成8—16个子核。随后细胞质纵裂,形成2、4、8或16个子原生质体,每个子原生质体分泌1层细胞壁,并生出两条鞭毛,子细胞由于母细胞壁胶化破裂而放出,长成新的植物体。
在某些环境下,如在潮湿的土壤上,原生质体再三分裂,产生数十、数百至数千个没有鞭毛的子细胞,埋在胶化的母细胞壁中,形成一个不定群体(palmella)。当环境适宜时,每个子细胞生出两条鞭毛,从胶质中放出(图1-38,B)。
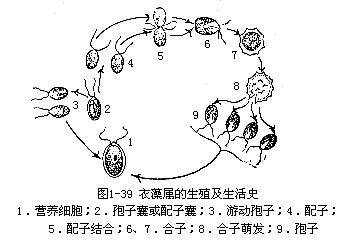
有性生殖 衣藻进行无性生殖多代后,再进行有性生殖。多数种的有性生殖为同配,生殖时,细胞内的原生质体经过分裂,形成32—64个小细胞,称配子。配子在形态上和游动孢子无大差别,只是比游动孢子小。成熟的配子从母细胞中放出后,游动不久,即成对结合,形成双倍、具四条鞭毛、能游动的合子。合子游动数小时后变圆,分泌形成厚壁合子,壁上有时有刺突。合子经过休眠,环境适宜时萌发,经过减数分裂,产生4个单倍核的原生质体,也有反复多次分裂,产生8—16—32个单倍核的原生质体。以后合子壁胶化破裂,单倍核的原生质体被放出,并在几分钟之内生出鞭毛,发育成新的个体(图1-39)。
衣藻大约有100多种,生活于含有机质的淡水沟和池塘中,早春和晚秋较多,常形成大片群落,使水变成绿色。
(二)团藻属(Volvox)
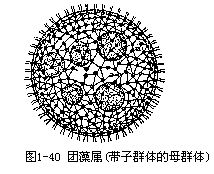
属于团藻目。植物体是由数百至上万个细胞,排列成1层空心球体,球体内充满胶质和水。细胞的形态和衣藻相同。每个细胞各有1层胶质包着,由于胶质膜彼此挤压,从表面上观察细胞为多边形的,各细胞间有原生质丝相连。群体后端有些细胞失去鞭毛,比普通营养细胞大十倍或十倍以上,称此为生殖胞(gonidium)。
无性生殖 由生殖胞进行多次纵分裂,形成皿状体(plakea),当皿状体发展为32个细胞时,细胞开始分化为营养细胞和生殖细胞,继续分裂直至形成1个球体,球体有1个孔。此时,群体内细胞的前端是向着群体中央,球体从孔经过翻转作用(inversion),细胞的前端翻转到群体的表面。翻转作用后,细胞长出两条鞭毛,子群体陷入母群体的胶质腔中。而后由母群体表面的裂口逸出,或待母群体破裂后放出(图1-40)。
有性生殖 为卵式生殖(cogamy)。群体中只有少数生殖细胞产生卵和精子。产生精子的生殖细胞,经过反复纵裂,形成皿状体,并经过翻转作用,发育成1个能游动的精子板(sperm packet)。游动精子板不分散成单个精子,而是整个精子板游近卵细胞附近才散开。产失卵的生殖细胞略膨大,不经分裂就发育成1个不动卵。精子穿过卵细胞周围的胶质,与卵结合形成合子。受精后,合子分泌出1个厚壁,厚壁可能是光滑的,也可能有刺状突起。合子从群体的胶质中放出后,不立即萌发,它能抵抗恶劣环境,数年不死,待环境好转时,即萌发。合子萌发前经过减数分裂,外壁层破裂,内壁层变成1个薄囊,原生质体在薄囊内发育成1个具有双鞭毛的游动孢子(或静孢子),游动孢子(或静孢子)连同薄囊一起,由外壁层的裂口逸出,发育成1个群体,其经过和无性生殖的次序相同(图1-41)。
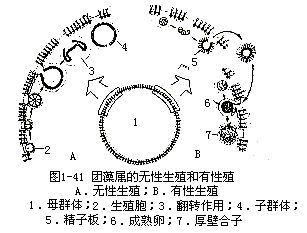
团藻经常在夏季发生于淡水池塘或临时性的积水中,二、三周后即消失。
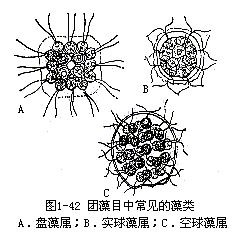
团藻目中常见的属有盘藻属(Gonium)、实球藻属(Pandorina)和空球藻属(Eudorina)(图1-42)。盘藻属是一种定形群体,无性生殖时,群体的全部细胞同时产生游动孢子,有性生殖为同配。实球藻属也是定形群体,无性生殖与盘藻属相同,有性生殖是异配。空球藻属是球形或椭圆形群体,少数种的群体细胞,有些是营养细胞,不产生配子和孢子,表明营养细胞和生殖细胞已开始有了分化,有性生殖为异配。从单细胞的衣藻属,群体的盘藻属、实球藻属、空球藻属和多细胞体的团藻属看,团藻目中有明显的演化趋势。藻体由单细胞、群体到多细胞体,细胞的营养作用和生殖作用,由不分工到分工,有性生殖由同配、异配到卵配3个方面演化。
(三)小球藻属(Chlorella)
小球藻属是色球藻目中的常见植物。植物体是单细胞浮游种类,细胞微小,圆形或略椭圆形,细胞壁薄,细胞内有1个杯形或曲带形载色体,细胞老熟时载色体分裂成数块。无蛋白核,只有蛋白核小球藻(Chlorella pyrenoidosa Chick.)有蛋白核。无性生殖时,原生质分裂形成2、4、8、16个似亲孢子,母细胞壁破裂时,孢子放出成为新的植物体(图1-43)。

小球藻在我国分布甚广,生活于含有机质的小河、沟渠、池塘等水中,在潮湿的土壤上也有分布。
(四)栅藻属(Scenedesmus)
栅藻属是绿球藻目中定型群体类型中的常见植物。一般是4个细胞的定形群体,也有8个或16个细胞的群体。细胞形状通常是椭圆形或纺锤形。细胞壁光滑或有各种突起,如乳头、纵行的肋、齿突或刺。细胞单核。幼细胞的载色体是纵行片状,老细胞则充满着载色体,有1个蛋白核。群体细胞是以长轴互相平行排列成1行,或互相交错排列成两行。群体中的细胞有同形或不同形的。无性生殖产生似亲孢子。产生似亲孢子时,细胞中的原生质体发生横裂,接着子原生质体纵裂,有的种连续发生一次或两次纵裂后,子原生质体变成似亲孢子,从母细胞壁纵裂的缝隙中放出,与纵轴相平行排列成子群体(图1-44)。
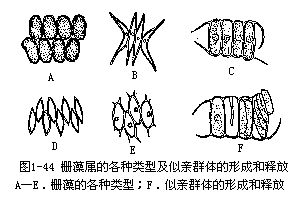
栅藻是淡水藻,在各种淡水水域中都能生活,分布极广。
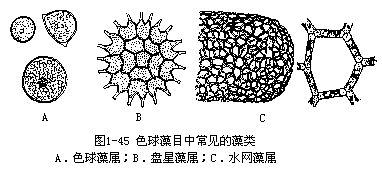
色球藻目中常见的单细胞类型有色球藻属(Chroococcus),是气生性藻类,生殖时产生游动孢子和配子。群体类型有盘星藻属(Pediastrum)和水网藻属(Hydrodictyon)。盘星藻属的群体由2—128个细胞组成,细胞排列成同心环状,外圈细胞的两角有突起。水网藻属是由许多长圆柱形的细胞连接成网状,每个网眼是由5—6个细胞组成(图1-45)。
(五)丝藻属(Ulothrix)

丝藻属是丝藻目中常见的植物。是单列细胞不分枝的丝状体。丝状体基部的细胞分化为有固着作用的细胞,叫固着器(holdfast)。固着器的载色体色较浅,小粒状。固着器之上为1列短筒形的营养细胞,细胞壁薄或厚,有层理,细胞单核位于中央,载色体是大形环带状。蛋白核多(图1-46)。无性生殖时,除固着细胞外,全部营养细胞均产生具4或2根鞭毛的游动孢子。1个细胞可产生2、4、8、16或32个游动孢子。每个子原生质体有1眼点,顶端有4根鞭毛,由母细胞侧壁的小孔放出。刚放出的游动孢子被1个薄囊包着,不久薄囊消失,游动孢子游动一段时间后,前端固着于基物上,产生细胞壁,再横分裂形成两个细胞,下面细胞为固着器,上面细胞继续分裂形成丝状体。有性生殖的配子产生过程和孢子形成过程相同,只是配子数目较多,可达64个。两个配子的结合是来自两条不同丝状体的同形配子的结合,称此种结合为异宗同配生殖(heterothallism)。合子经过休眠及减数分裂,产生游动孢子或静孢子,每个孢子长成1个新植物体。无性生殖或有性生殖可同时在1条丝状体上进行。丝藻属有的种类可通过丝状体的断裂,进行营养繁殖。
丝藻属多生活于流动的淡水中,在瀑布或急流水的岩石上较多,湖泊的岸边也可采到。常丛生在一起,呈矮的绿色绒毯状。
(六)石莼属(Ulva)
石莼属是石莼目植物。植物体是大形的多细胞片状体,由两层细胞构成。片状体呈椭圆形、披针形、带状等。植物体下部长出无色的假根丝,假根丝生在两层细胞之间并向下生长,伸出植物体外,互相紧密交织,构成假薄壁组织状的固着器,固着于岩石上。固着器是多年生的,每年春季长出新的植物体。藻体细胞排列不规则但紧密,细胞间隙富有胶质。表面观,细胞为多角形;切面观,细胞为长形或方形。细胞单核,定于片状体细胞的内侧。载色体片状,位于片状体细胞的外侧,有1枚蛋白核(图1-47)。

石莼属有两种植物体,即孢子体(sporophyte)和配子体(gametophyte)。两种植物体都是两层细胞的植物。成熟的孢子体,除基部外,全部细胞均可形成孢子囊。孢子母细胞经减数分裂,形成单倍体的游动孢子,游动孢子具4根鞭毛。孢子成熟后脱离母体,游动一段时间后,附着在岩石上,约二、三天后萌发成配子体,此期为无性生殖。成熟的配子体产生许多同型配子。配子的产生过程和孢子相似,但产生配子时,配子不经过减数分裂。配子具两根鞭毛。配子结合是异宗同配。合子二、三天后即萌发成孢子体。此期为有性生殖。在石莼属的生活史中,就核相来说,从游动孢子开始,经配子体到配子结合前,细胞中的染色体是单倍的,称配子体世代(gametophyte generation)或有性世代(sexual generation)。从结合的合子起,经过孢子体到孢子母细胞止,细胞中的染色体是双倍的,称孢子体世代(sporophyte generation)或无性世代(asexual generation)。二倍体的孢子体世代和单倍体的配子体世代互相更替,称为世代交替(alternation of generations)。在形态构造上基本相同的两种植物体,互相交替循环的生活史,叫同形世代交替(isomorphic alternation of generations)。
石莼目中的浒苔属(Enteromorpha)和礁膜属(Monostroma)与石莼属相近。浒苔属是1层细胞的管状体,生活史与石莼属相同。礁膜属是1层细胞膜状体,生活史是异形世代交替(heteromorphic alternation of generations)(图1-48)。

(七)刚毛藻属(Cladophora)
属于刚毛藻目。植物体是分枝的丝状体。以基细胞固着于基质上。细胞长圆柱形,细胞壁厚,分3层,内层为纤维素,中层为果胶质,外层是1种不溶性物质,有人认为是几丁质。细胞中央有1个大液泡,载色体网状,壁生,含有多数蛋白核。细胞多核。细胞分裂时,细胞侧壁的中部生出1个环,此环向中央生长,将细胞隔成两个。分枝是从1个细胞顶端的侧面发生,使分枝常常像二叉状。分枝一般在靠近丝状体顶端的一些细胞里发生(图1-49)。
刚毛藻属中有些种是同形世代交替,有些是异形世代交替。在形态构造上显著不同的两种植物体,互相交替循环的生活史,叫异形世代交替。
刚毛藻属约有150种,分布很广,淡水、海水都产。藻体常常大量繁殖,可以制成食品。
(八)松藻属(Codium)
属于管藻目。全部海产,固着生活于海边岩石上。植物体是管状分枝的多核体,许多管状分枝互相交织,形成有一定形状的大型藻体,外观叉状分枝,似鹿角,基部为垫状固着器。丝状体有一定分化,中央部分的丝状体细,无色,排列疏松,无一定次序,称作髓部,向四周发出侧生膨大的棒状短枝,叫做胞囊(utricle),胞囊紧密排列成皮层部。载色体数多,小盘状,多分布在胞囊远轴端部分,无蛋白核。细胞核极多而小。髓部丝状体的壁上,常发生内向生长的环状加厚层,有时可使管腔阻塞,其作用是增加支持力,这种加厚层在髓部丝状体上各处都有,而胞囊基部较多。
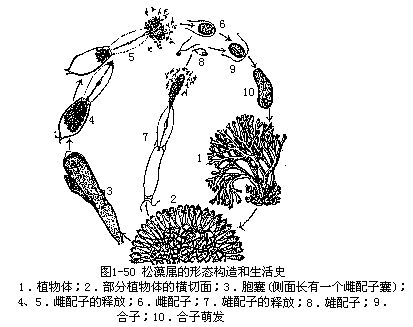
松藻属植物体是二倍体。有性生殖时在同一藻体或不同藻体上生出雄配子囊(malegametangium)和雌配子囊(female gametangium)。配子囊发生于胞囊的侧面,配子囊内的细胞核一部分退化,一部分增大。每个增大的核经过减数分裂,形成4个子核,每个子核连同周围的原生质一起,发育成具双鞭毛的配子。雌配子大,含多个载色体;雄配子小于雌配子数倍,只含有1—2个载色体。配子结合为合子后,立即萌发,长成新的二倍体植物(图1-50)。
(九)水绵属(Spirogyra)
水绵属是接合藻目中的常见植物。植物体是由1列细胞构成的不分枝的丝状体,细胞圆柱形。细胞壁分两层,内层为纤维素构成,外层为果胶质。壁内有1薄层原生质,载色体带状,1至多条,螺旋状绕于细胞周围的原生质中,有多数的蛋白核纵列于载色体上。细胞中有大液泡,占据细胞腔内的较大空间。细胞单核,位于细胞中央,被浓厚的原生质包围着。核周围的原生质与细胞腔周围的原失质之间,有原生质丝相连(图1-51)。

接合生殖 水绵属的有性生殖多发生在春季或秋季。生殖时两条丝状体乎行靠近,在两细胞相对的一侧相互发生突起,突起渐伸长而接触,于是接触的壁消失,连接成管,称为接合管(conjugation tube)。同时,细胞内的原生质体放出一部分水分,收缩形成配子。第一条丝状体细胞中的配子,以变形虫式的运动,通过接合管移至相对的第二条丝状体的细胞中,并与细胞中的配子结合。结合后,第一条丝状体的细胞只剩下1条空壁,此种丝状体是雄性的,其中的配子是雄配子;而第二条丝状体的细胞在结合后,每个细胞中都有1个合子,此种丝状体是雌性的,其中的配子是雌配子。配子融合时细胞质先融合,稍后两核才融合形成接合子。两条接合的丝状体和它们所形成的接合管,外观同梯子一样,这种接合叫梯形接合(scalariformconjugation)(图1-51,C)。合子成熟时分泌厚壁,分3层:内层薄,是纤维质;中层厚,是纤维质并稍带几丁质;外层薄,是纤维质或果胶质。合子内充满贮藏物质,初期是淀粉,后期转变为脂肪。成熟的合子随着死亡的母体沉于水底,待母体细胞破裂后放出体外。有些种类则进行侧面接合(lateral conjugation),侧面接合是在同1条丝状体上相邻的两个细胞间形成接合管,或两个细胞之间的横壁上开1孔道,其中1个细胞的原生质体通过接合管或孔道移入另1个细胞中,与其细胞中的原生质融合形成合子(图1-51,B)。侧面接合后,丝状体上空的细胞和具合子的细胞,是交替存在于同1条丝状体上,这种水绵可以认为是雌雄同体的。梯形接合与侧面接合比较,侧面接合较为原始。
合子耐旱性很强,水涸不死,待环境适宜时萌发。一般是在合子形成后数周或数月,甚至一年以后萌发。萌发时,核先减数分裂,形成4个单倍核,其中3个消失,只有1个核萌发,形成萌发管,由此长成新的植物体(图1-51,D)。
水绵属全部是淡水产,是常见的淡水绿藻,在小河、池塘、沟渠或水田等处均可见到,繁盛时大片生于水底或成大块漂浮水面,用手触及有粘滑的感觉,这点是本目丝状藻和其他丝状藻类显著不同的地方。
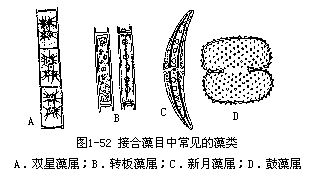
接合藻目中常见的丝状藻类有双星藻属(Zygnema)和转板藻属(Mougeotia)。单细胞类型的有新月藻属(Closterium)和鼓藻属(Cosmarium)(图1-52)。
(十)轮藻属(Chara)
属于轮藻纲轮藻目植物。植物体直立,具分枝,体表常含有钙质,以单列细胞分枝的假根固着于水底淤泥中。主枝分化成节和节间,节的四周轮生有短枝,短枝也分化成节和节间,短枝又被叫做“叶”。无论是主枝或是短枝,顶端都有1个半球形细胞,叫做顶端细胞(apicacell),植物的生长即由顶端细胞不断分裂形成的。顶端细胞横分裂,形成两个子细胞,上面的子细胞继续保持顶端细胞的作用(图1-53)。下面的子细胞再进行一次横分裂,又形成两个子细胞。第二次形成的两个子细胞中,下面的1个不再分裂,长大成节间的中央细胞;上面的1个经过数次纵裂,构成节部,同时,部分细胞发育成包围于间节中央细胞外的皮层。节部细胞短小,节间细胞长管状、多核、载色体多数,具中央大液泡。主枝能无限生长;短枝到一定长度便停止生长,在短枝节上还具有单细胞刺状突起。
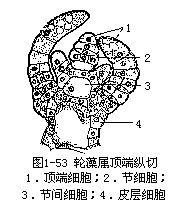
轮藻属的有性生殖是卵式生殖。雌性生殖器官叫卵囊(oogonium),雄性生殖器官叫精子囊(spermatangium),雌雄生殖器官皆生于短枝的节上。卵囊生于刺状体上方,长卵形,内含1个卵细胞,卵的外围有5个螺旋状的管细胞(tube cell)。管细胞上有1个小细胞,叫冠细胞(corona cell),5个冠细胞在卵囊上组成冠(corona)。精子囊生于刺状体下方,圆形,外围有8个三角形细胞,叫盾细胞(shield cell)。盾细胞内含有很多桔红色的载色体,因此,成熟的精子囊,肉眼观看是桔红色的。盾细胞内侧中央连接1个圆柱形细胞,叫盾柄细胞(manubrium)。盾柄细胞末端有1-2个圆形细胞,叫头细胞(head cell)。头细胞上又可生几个小圆细胞,叫次级头细胞。从这些次级头细胞上,长出多条单列细胞的精囊丝(antheridial filament),每个细胞内产生1个精子,成熟时,精子释放到水中。精子细长,顶端生两个等长鞭毛。卵囊成熟时,冠细胞裂开,精子从裂缝进入,与卵结合。合子分泌形成厚壁(图1-54)。
合子经过休眠后萌发,萌发时合子核分裂,形成4个子核,而后继续发育成原丝体,有节和节间的分化,由原丝体上可长出数个新植物体。有人认为,合子第一次分裂形成4个子核是减数分裂。
轮藻常常以藻体断裂的方式进行营养繁殖,断裂的藻体沉在水底,长出假根和芽,发育成新的植物体。轮藻体基部可长出珠芽,由珠芽长出植物体。珠芽含有大量淀粉,类似种子植物的块根或块茎。
轮藻目常见的丽藻属(Nitella),与轮藻属的区别有两点:一是每个管细胞上面有两个冠细胞,整个卵囊上有10个冠细胞,分两层,每层5个;其次是节间无皮层(图1-55)。
轮藻的植物体高度分化,生殖器官构造复杂,外面有1层营养细胞包着,可与高等植物的性器官比较,细胞有丝分裂与陆生绿色植物相似,因此,有人将它们列为独立1门。
轮藻多生于淡水,在不大流动或静水的底部大片生长,少数生长在微盐性的水中。

三、绿藻门在植物界中的地位
绿藻化石很多,比较早的化石大约发现在14—12亿年前。在我国河北省蓟县震旦亚界地层中,发现了一种12亿年前的真核多核体藻类,个体中央有中轴,两侧有许多轮状排列的侧枝,经鉴定属于绿藻纲管藻目多毛藻科的真核生物,定名为震旦塔藻化石。
1975年,美国藻类学家柳文,在一种海鞘的泄殖腔沟纹处,发现了一种具有叶绿素a和b的原核藻类,并定名为原绿藻。之后,不少人认为,真核绿藻是由原核的原绿藻演化而来的,人们把原绿藻的发现看作是藻类进化史上的一件大事,称原绿藻为“活化石”。
绿藻和高等植物之间有很多相似之处,它们有相同的光合作用色素,光合作用产物都是淀粉,鞭毛类型都是尾鞭型。因此,多数植物学工作者承认高等植物的祖先是绿藻。绿藻门在植物界的系统发育中,居于主干地位。然而高等植物究竟是从哪一类绿藻发展来的,还没有直接答案。有些学者主张高等植物起源于绿藻中的轮藻,因为轮藻的色素、贮藏食物和鞭毛结构与高等植物相同。藻体结构和生殖器官的构造,与高等植物比较相近;合子萌发时产生的原丝体和苔藓植物近似。但是,他们忽略了一个重要事实,轮藻合子萌发时为减数分裂,不形成二倍体的营养体,没有孢子行无性生殖,所以高等植物不能起源于轮藻。然而高等植物到底起源于哪种绿藻,现在还没有可靠的证据。在印度、非洲和日本发现的费氏藻(Fritshiella tuberosaIyengar)有直立枝和匍匐枝的分化,匍匐枝生于地下,直立枝穿过薄土层,在土表分成丛状枝,外表有角质层,有世代交替现象,能适应陆地生活。鲍尔(Bower,1935)认为,高等陆生植物可能是从古代这种类型的绿藻发展来的。
免责声明:本文来源于网络,文中有些文字或数据已经过期失效,仅供学习备课参考!
电脑版地址:http://www.cgzdl.com/shuku/244/9723.html
手机版地址:http://m.cgzdl.com/shuku/244/9723.html
-lp.jpg "甘肃省兰州市旅游地图高清版")






